 Cell Technology Co., Ltd.")

News
News Center
Attention, working professionals: Lack of sleep can seriously shorten your lifespan.
2021-04-01
Some say that there’s no such thing as "easy" in the life of a working professional—things like falling asleep easily or staying asleep can be real challenges. In fact, data shows that 45.4% of people experienced insomnia to varying degrees over the past year.
Anyone who’s been there knows how agonizing insomnia can be— the harder you try to fall asleep, the more elusive sleep becomes...

Sleep deprivation initially manifests as fatigue, weakness, irritability, and difficulty concentrating. Over the long term, chronic sleep loss can also lead to obesity, heart disease, high blood pressure, and diabetes—and may even shorten a person’s lifespan.
Severe sleep deprivation can actually "kill" you.
A study titled "Sleep Loss Can Cause Death through Accumulation of Reactive Oxygen Species in the Gut," published in the international journal *Cell*, concluded that severe sleep deprivation can lead to death, according to Dr. Alexandra Vaccaro and Dragana Rogulja from Harvard University's Department of Neurobiology. [1] 。

Research has found that sleep deprivation in flies and mice leads to death caused by oxidative stress, with the gut playing a central role in this process. Further studies also revealed that survival without sleep becomes possible if the buildup of reactive oxygen species (ROS) is prevented—as illustrated in Figure 1.
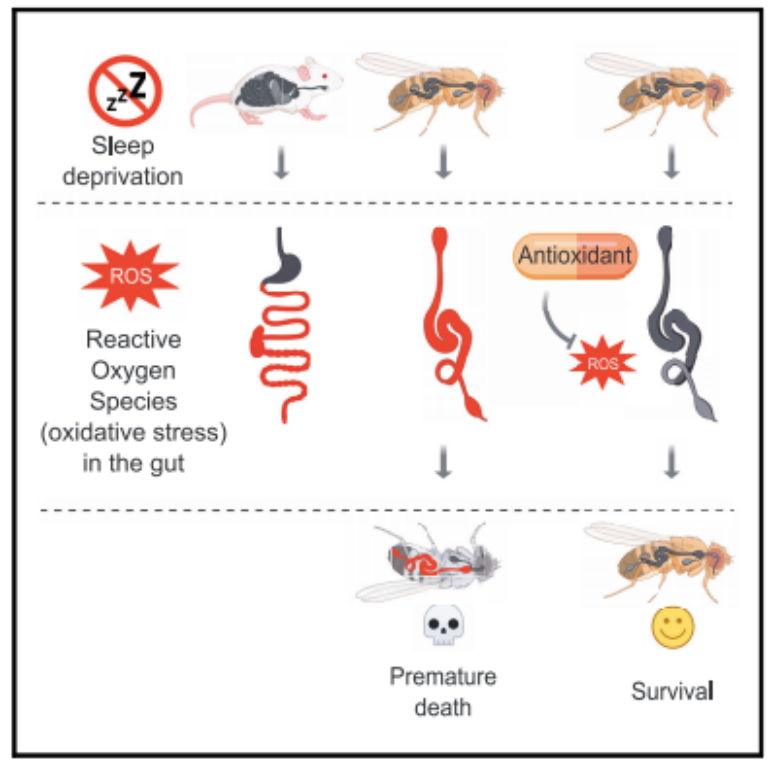
Figure 1: Sleep deprivation leads to the accumulation of reactive oxygen species (ROS) in the guts of flies and mice, triggering oxidative stress responses in their organs. However, when ROS accumulation is prevented using antioxidants, flies can survive normally—even without sleep.
Dr. Alexandra Vaccaro’s lab team used flies and mice as experimental subjects because these insects and mammals share fundamental sleep-related characteristics, and sleep is essential for the survival of flies. To investigate the effects of chronic sleep deprivation, Dr. Vaccaro’s team employed gene-editing techniques to systematically reduce the animals’ sleep duration—without eliminating their innate physiological need for sleep. Surprisingly, they discovered that after genetically restricting sleep, reactive oxygen species (ROS) accumulated significantly in the gut.
Gal4 is a gene transcription activator that targets neurons, and its activation leads to severe sleep deprivation. Dr. Alexandra Vaccaro’s team successfully used Gal4-driven gene expression to enable sleep-regulating neurons to express the temperature-sensitive channel protein TrpA1. At 21°C, TrpA1 remains inactive—this temperature serves as a baseline for monitoring normal sleep patterns. However, when the temperature is raised to 29°C, TrpA1 opens, triggering neuronal activity and significantly reducing sleep in the animals. Specifically, when TrpA1 was expressed in neurons labeled by either 11H05-Gal4 or 60D04-Gal4 (11H05>TrpA1, 60D04>TrpA1), flies experienced a dramatic 90% reduction in sleep (Figures 2A–D).
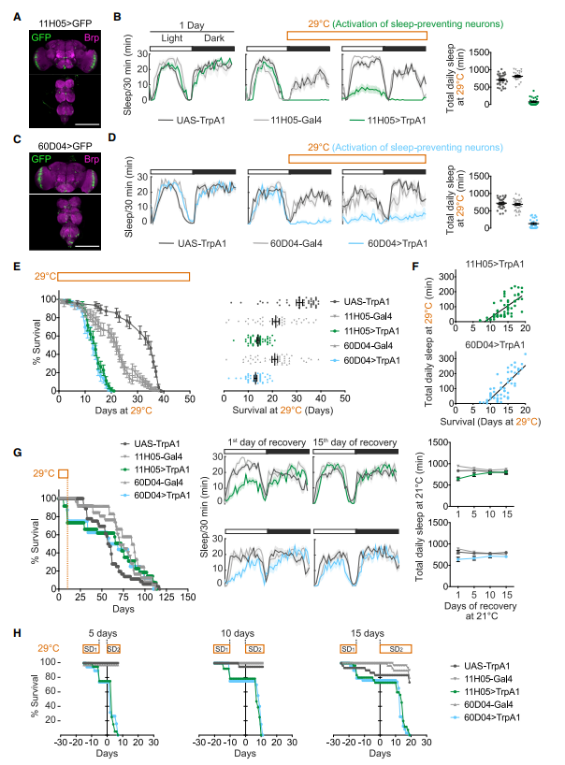
Figure 2: Sleep deprivation can shorten lifespan.
A. Expression of 11H05-Gal4 in the nervous system: Top image—brain; bottom image—ventral nerve cord. Brp is a presynaptic protein that labels neuronal fibers.
B. Day 1, 21C: The control group showed no difference in sleep duration compared to the two experimental groups—flies expressing TrpA1 under the 11H05-Gal4 driver (11H05>TrpA1) and flies with TrpA1 expression alone. Male flies exhibited typical diurnal and nocturnal sleep patterns. However, on Day 2 (marked in orange), when the temperature was raised to 29°C, the 11H05>TrpA1 flies virtually lost their ability to sleep. Right panel: Daily sleep patterns of the control and experimental fly groups.
C and D are the same as A and B, but this is more pronounced in 60D04-Gal4.
E. Survival rates of sleep-deprived (green, blue) and non-deprived fruit flies at 29°C. Left panel: Population; right panel: Individual survival.
F. The correlation between individual sleep patterns and lifespan.
G. Left panel: Survival rate of flies able to sleep after being deprived for 10 days. Middle-right panel: Sleeping during the recovery period.
H. The flies were deprived of sleep for 10 days (SD1) and then allowed to sleep for 5, 10, or 15 days before being deprived again (SD2). On day 0, SD2 onset occurred.
Researchers found that, under all conditions, sleep-deprived animals died earlier than their counterparts in the control group who were not deprived of sleep. On the tenth day of sleep deprivation, the animals exhibited the highest mortality rate. Upon examining these animals, researchers observed that most of their tissues appeared unaffected—but notably, levels of reactive oxygen species (ROS) in the intestines of the sleep-deprived animals had risen significantly.
To detect the levels of ROS in the gut and other organs, researchers used dihydroethidium, which emits red fluorescence upon oxidation but blue fluorescence when unoxidized. Meanwhile, another detection method—activated by hydrogen peroxide—also confirmed the increased presence of ROS in the intestinal tissue.
Afterward, the researchers tested whether this conclusion holds true across mammalian models. They used mild, continuous mechanical stimulation to keep C57BL/6J mice sleep-deprived for up to 5 days. In each repeated experiment, 5 mice were placed in a specialized device equipped with a slowly rotating bar, preventing them from falling asleep. As a control group, another 5 mice were housed in the same apparatus—but without the rotating bar.
Researchers simultaneously examined the internal organs of mice both with and without sleep deprivation. They sliced the tissues and then used DHE (dihydroethidium) to detect signs of ROS. Compared to mice that had adequate sleep, those lacking sleep showed increased levels of ROS in both the small and large intestines—particularly in the small intestine, where ROS levels spiked after just two days of sleep deprivation. In contrast, the rise in ROS within the large intestine (colon) was less pronounced, likely because the colon already had higher baseline levels of ROS. Importantly, no changes in ROS levels were observed in other organs of the mice—even after five days of sleep deprivation. (As shown in Figure 3.)
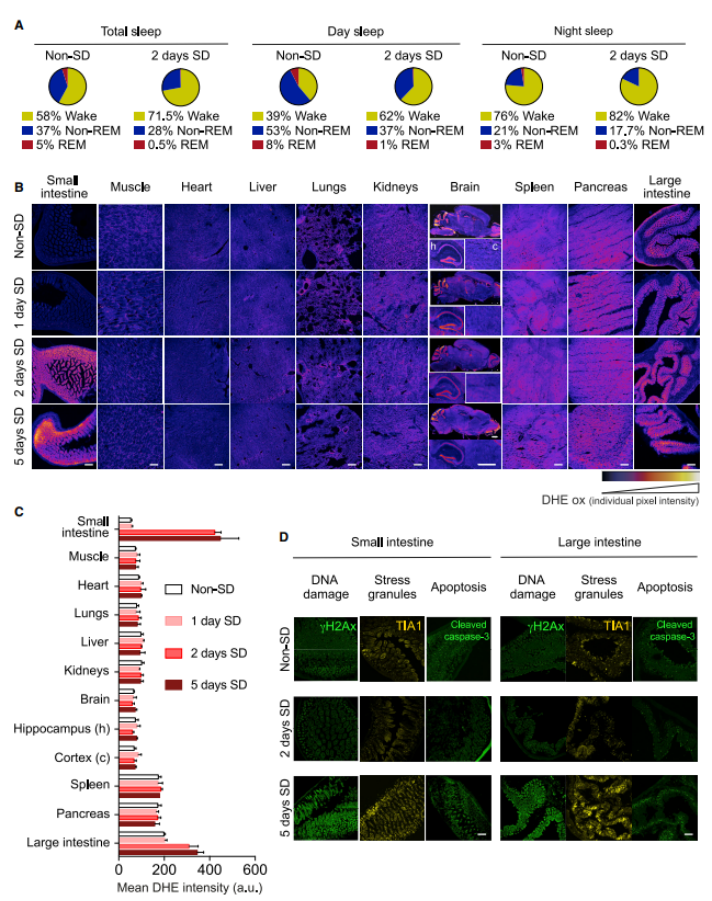
Figure 3: ROS Accumulation in the Mouse Intestine During Sleep Deprivation
A. EEG/EMG recordings of REM sleep, non-REM sleep, and wakefulness in C57BL/6J mice subjected to sleep deprivation (SD) and those maintained under non-deprived (non-SD) conditions.
B. After just two days of sleep deprivation, levels of ROS in the small and large intestines were elevated.
C. Quantitative analysis of ROS levels in tissues from sleep-deprived and non-deprived mice.
D. Oxidative stress markers in the small and large intestines only increase after 5 days of sleep deprivation.
The results from the two sets of experiments revealed the link between sleep deprivation, ROS accumulation, and animal mortality. To establish a causal relationship among these factors, Dr. Alexandra Vaccaro’s team further investigated whether it’s possible to extend an animal’s lifespan by eliminating ROS—without necessarily increasing sleep duration.
Researchers tested 53 known substances with antioxidant properties through dietary supplementation and discovered that 11 of them enabled sleep-deprived animals to maintain a normal or near-normal lifespan. Among these antioxidants, some directly neutralize ROS by donating electrons (such as lipoic acid, quercetin, and 4-hydroxy-2,2,6,6-tetramethylpiperidine-1-oxyl), while others reduce ROS levels by boosting the expression of naturally occurring antioxidant enzymes (e.g., N-acetylcysteine and sodium phenylbutyrate). Notably, despite the continuous use of these antioxidants, the animals remained sleep-deprived—meaning these compounds did not work by simply extending their sleep duration. (As shown in Figure 4.)
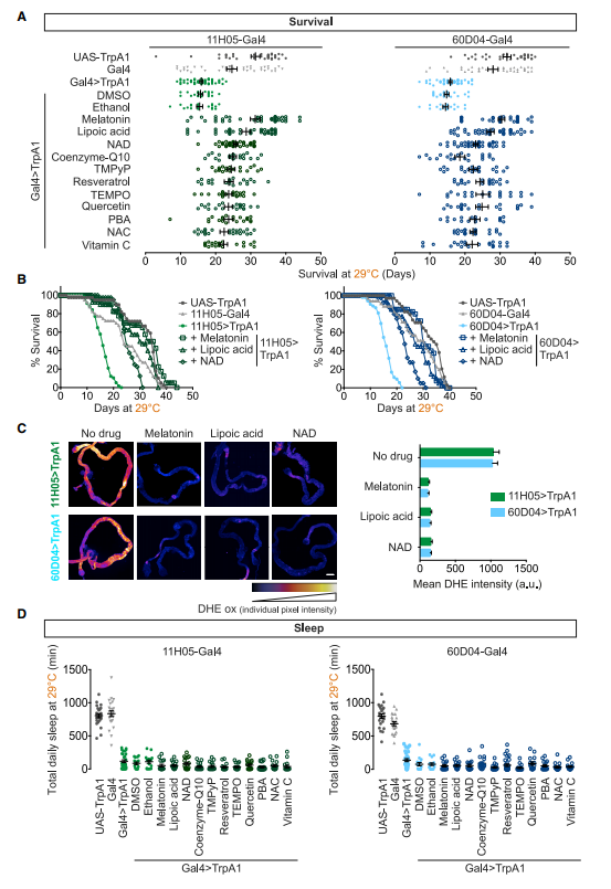
Figure 4: Eliminating ROS prevents the death of heat-stressed sleep-deprived fruit flies.
A. Individual survival: Sleep-deprived fruit flies (11H05>TrpA1, 60D04>TrpA1) died earlier than the control group (11H05-Gal4, 60D04-Gal4, UAS-TrpA1). Neither the solvent (ethanol nor dimethyl sulfoxide) affected survival. However, supplementing with antioxidants (melatonin + vitamin C) significantly extended the lifespan of sleep-deprived fruit flies.
B. Example survival curves for the three rescue compounds.
C. Reactive oxygen species are cleared from the gut by survival-promoting compounds. Representative images on day 10.
D. Rescue antioxidants do not promote sleep.
Embrace health—start with the dynamic balance of ROS.
ROS are the primary mediators through which phagocytes carry out their phagocytic and cytotoxic functions; however, under pathological conditions, when reactive oxygen species production exceeds the body's capacity to neutralize them, the organism triggers an oxidative stress response, leading to oxidative damage. In essence, the accumulation of these ROS causes DNA damage, accelerating aging, injury, apoptosis, and even mutations—or even cancer—in cells (including stem cells), ultimately setting off a vicious cycle. [2-5] 。
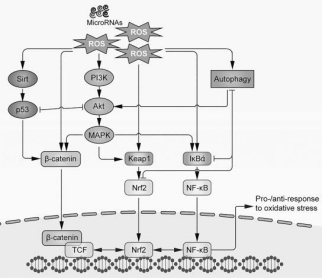
Figure 5: Signaling pathways involved in oxidative stress-induced stem cell senescence.
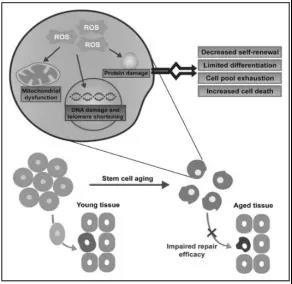
Figure 6: Aging stem cells lose their regenerative and differentiation capabilities, leading to the depletion of the functional stem cell pool and severely impairing their ability to repair damaged tissues.
In cell biology, the redox environment regulates numerous physiological and pathological processes. Oxidative stress disrupts the body's redox balance, simultaneously triggering stem cell aging and severely impacting the internal milieu as well as tissue repair mechanisms.
The consequences of stem cell aging are both subtle and pronounced, ranging from disruptions in cellular homeostasis to increased cellular damage, loss of regenerative capacity, and heightened cell death. This process is intricately regulated by a multitude of intrinsic and extrinsic pathways. Maintaining ROS at an optimal physiological level within cells—rather than simply reducing them altogether—is crucial for preserving redox balance, which plays a vital role in disease prevention and delaying the aging process. Research clearly highlights the critical role of oxidative stress in triggering stem cell senescence, while also revealing how stem cells, through their robust paracrine signaling, can modulate endogenous antioxidant defenses to sustain cellular homeostasis.
MSCs exhibit high resistance to oxidative damage, a fact that has been confirmed in numerous studies. For instance, bone marrow-derived mesenchymal stem cells cultured in vitro demonstrate resilience against oxidative and nitrosative stress, which is closely linked to their robust expression of antioxidant enzymes such as SOD1, SOD2, catalase (CAT), glutathione peroxidase (GPx), and the antioxidant molecule glutathione (GSH). [6] The lack of GSH leads to a loss of tolerance to oxidative stress. [7] SIRT6 is believed to enhance MSCs' resistance to oxidative damage and ROS production by upregulating downstream antioxidants, including heme oxygenase-1 (HO-1). [8] Etc.
MSCs' ability to modulate these processes can explain the diversity of MSC-based therapies in disease models, as well as the protective effects of MSCs across species. On April 20, 2020, a research team from the University of Melbourne in Australia introduced a new concept regarding the mechanism of action of mesenchymal stem cells—specifically, their antioxidant function—in an article published in the journal *Stem Cell Translational Medicine*. [9] , as illustrated in Figure 7, its mechanism of action is well-documented. Stem cell supplementation, recognized as a promising target for preventing age-related decline in the body, has been attracting increasing attention.
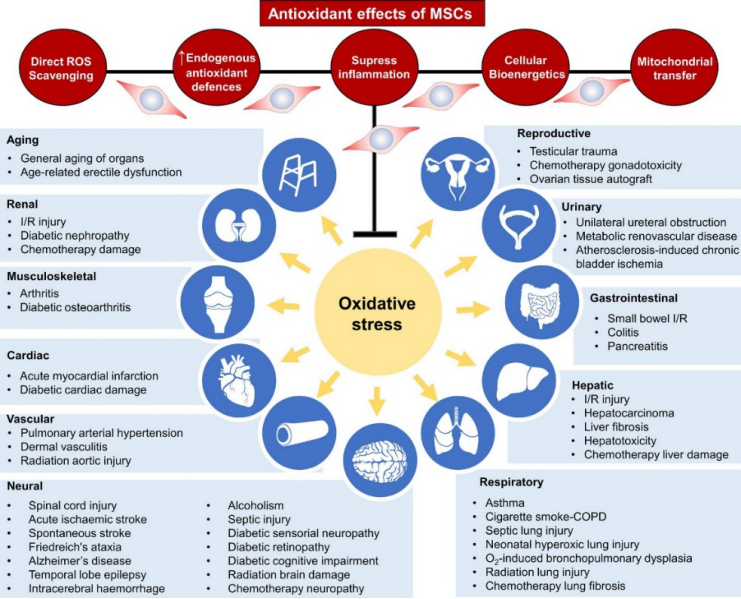
Figure 7: Antioxidant Effects of Bone Marrow Mesenchymal Stem Cells.
The image shows that bone marrow mesenchymal stem cells directly scavenge reactive oxygen species, upregulate intrinsic antioxidant defenses, suppress inflammation, and reprogram cellular bioenergetics—all while donating mitochondria to enhance their antioxidant properties. Meanwhile, the immunosuppressive capabilities of MSCs help prevent excessive ROS production. Together, these mechanisms reduce oxidative stress and maintain physiological levels of ROS in the body. Importantly, the antioxidant effects of bone marrow mesenchymal stem cells are directly linked to therapeutic benefits in a range of diseases as well as anti-aging applications.
Jiuzhitang Maker's U.S. partner, Stemedica, has developed its clinically validated cell-production platform, BioSmart™, at its cGMP-compliant facility in California. This platform mimics the natural microenvironment of cells within the human body and is designed to produce ischemia-tolerant human bone marrow mesenchymal stem cells (it-hMSCs) under continuous low-oxygen conditions—resulting in cells with enhanced resilience. These advanced cells are expected to play a more significant role in anti-aging therapies and treatments for insomnia.
1. Hypoxic culture, Stronger cloning ability
2. Hypoxic culture, Has a stronger chondrogenic differentiation ability

3. Hypoxic Culture , which enhances gap junction formation
4. Hypoxic culture, Enhanced chemotactic homing ability
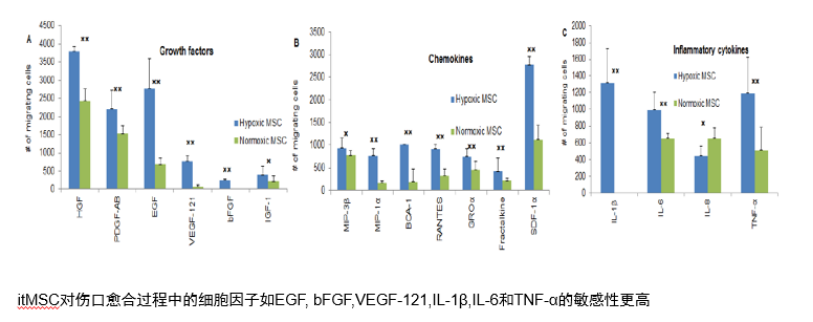
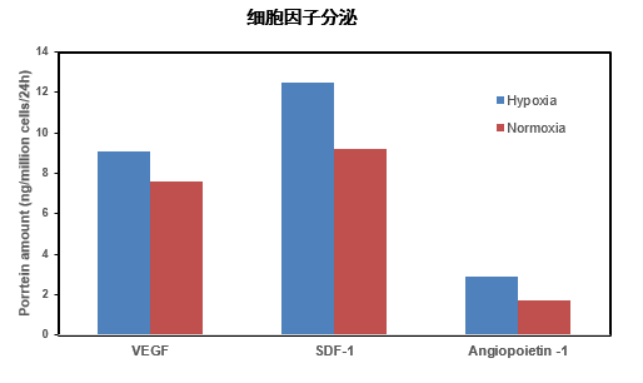
5. Hypoxic culture, Has stronger paracrine ability
People often feel both curiosity and fear toward new developments. Bone marrow mesenchymal stem cells have gone from being questioned in the past to gradually gaining acceptance—and now, they’re experiencing a remarkable boom. Their value is bound to reach new heights, positioning them as a powerful biological resource at the heart of the global health industry.
References:
1. Sleep Loss Can Lead to Death Through the Accumulation of Reactive Oxygen Species in the Gut
2. Imbalanced Production of Reactive Oxygen Species and Mitochondrial Antioxidant SOD2 in Fabry Disease-Specific Human Induced Pluripotent Stem Cell-Differentiated Vascular Endothelial Cells
3. Repeated Mild Traumatic Brain Injury: Potential Mechanisms of Damage
4. Cell Contact Accelerates Replicative Senescence of Human Mesenchymal Stem Cells Independent of Telomere Shortening and p53 Activation: The Roles of Ras and Oxidative Stress
5. Production of Reactive Oxygen Species by Multipotent Stromal Cells/Mesenchymal Stem Cells Upon Exposure to Fas Ligand
6. Induced Pluripotent Stem Cells Without c-Myc Alleviate Acute Kidney Injury by Downregulating Oxidative Stress and Inflammation Signaling in Ischemia-Reperfusion Rats
7. Chen M-F, Lin C-T, Chen W-C, et al. Sensitivity of human mesenchymal stem cells to ionizing radiation. Intern J Radiat Oncol Biol Phys. 2006;66(1):244-253.
8. Valle-Prieto A, Conget PA. Human mesenchymal stem cells efficiently manage oxidative stress. Stem Cells Dev. 2010;19(12):1885-1893.
9. Liu T, Ma X, Ouyang T, et al. SIRT1 reverses cellular senescence by enhancing autophagy and attenuates oxidative stress-induced apoptosis through promoting p53 degradation. Int J Biol Macromol. 2018;117:225-234.
Related News

Here is the title—h1 placeholder text


Copyright © Jiuzhitang Maker (Beijing) Cell Technology Co., Ltd.
Powered by: 300.cn SEO | Privacy Policy